Вступ.
При деполяризації мембрани багато потенціал-залежних йонних каналів, особливо деякі К+ канали переходять в довготривалий стан непровідності, чи інактивації. В результаті цього істотно змінюється транзієнтна проникність мембрани для калію, не зважаючи на тривалу і сильну деполяризацію. Така властивість деяких К+ каналів дозволяє дуже тонко регулювати палітру активності збудливих клітин через різну частоту потенціалів дії. Тому, вочевидь, дуже важливо для біологів та біомембранологів розуміти молекулярні механізми, які викликають інактивацію К+ каналів та можливі способи фізіологічної регуляції цього процесу.
Інактивація К+ каналів
|
|
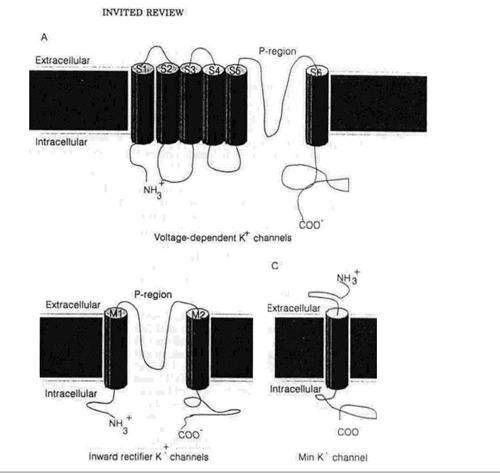
На відміну від гомогенної інактиваційної поведінки потенціал-залежних Na+ каналів, потенціал-залежні К+ канали, досліджені в їхньому природному середовищі, показують широку різноманітність інактиваційних часових інтервалів. Інактивація цих каналів спостерігається в діапазоні від кількох мілісекунд – швидка інактивація (як у А-струму, описаного в нейронах молюсків) до майже помітного процесу тривалістю кілька тисяч мілісекунд, (як у випадку струму затриманого випрямлення в гігантському аксоні кальмара). Така різноманітність також спостерігається в дослідах з клонованими К+ каналами, експресованими в гетерогенному середовищі. Наприклад, ShakerB канал інактивується з часовою константою 1-3 мс, тоді як відповідний канал drk1 демонструє значно повільнішу інактивацію. Так само, К+ канали, клоновані з мозку ссавців, показують дуже відмінні інактиваційні інтервали. В деяких випадках неструктурні білки, що утворюють minК+ канали (мал.1) не піддаються інактивації навіть після деполяризаційних імпульсів тривалістю кілька секунд. Ця різноманітність інактиваційних інтервалів навіть між дуже близькими між собою продуктами генів допомогла краще зрозуміти механізми інактивації К+ каналів.
NH2 – кінцевий домен як інактиваційної частинки.
Відомо, що Shaker канали - це родина різних, але близько споріднених білків, які кодуються транскриптами альтернативного сплайсингу. Існує 5 альтернативно сплайсованих NH2 – кінцевих варіантів Shaker і 2 СООН – кінцевих. Після того, як вдалося встановити продукт Shaker локусу дрозофіли, дослідники припустили, що альтернативні транскрипти одного й того ж самого локусу індукують експресію К+ каналів з різною інактиваційною поведінкою.
Таким чином, наприклад, струм через ShakerА2 – канал, який відрізняється від ShakerD2 лише своїм NH2 – кінцевим доменом, на 90% інактивується протягом кількох мілісекунд, тоді як струм через ShakerD2 канал інактивується лише на 15% і значно повільніше.
Iverson and Rudy створювали конструкції з різними комбінаціями NH2 – та СООН – кінцевих варіантів Shaker і помітили виникнення як інактиваційних, так і неінактиваційних струмів після їхньої експресії. Помічена різниця може бути віднесена на рахунок присутності саме NH2 – кінцевих доменів в кожній конструкції. Ці результати дозволили твердити про те, що структури, розташовані на NH2– кінці (можливо це цитоплазматичні домени) можуть бути задіяні в інактиваційному процесі.
Працюючи з ShakerВ каналами Hoshi та ін. простежили, що обробка трипсином внутрішнього боку збудливої мембрани катастрофічно сповільнює інактивацію ізольованих каналів, не впливаючи на процеси активації та проникності. Ці дані узгоджуються з припущенням про участь цитоплазматичного домену в процесі інактивації і через це вчені зосередили свою увагу на аналізі амінокислотної послідовності кінцевого NH2 домену ShakerВ білку. Hoshi та співробітники робили систематичні делеції амінокислот і встановили, що делеції, які включають перші 22 амінокислоти змінюють швидкий інактиваційний процес на зразок як це робить трипсин. Такі делеції збільшують середній час, протягом якого канали перебувають у відкритому стані, збільшують кількість відкритих протягом серії каналів та зменшують тривалість інтервалів між серіями. Ці дані відповідають результатам про сповільнення інактивації макроскопічних струмів. Делеції поза ділянкою перших 22 амінокислот не порушують інактивацію, але зменшують середню тривалість відкритості каналів пропорційно до довжини делетованої ділянки. Ці спостереження доводять роль перших 22 амінокислот як інактиваційної “кульки”, а решти NH2 кінцевого домену як “ланцюжка”, що сполучає кульку з каналом.
Прогноз про те, що, виходячи з моделі “кульки і ланцюжка” додавання ізольованих кульок до цитоплазматичного боку каналів, не здатних до інактивації відновить цю їхню здатність підтвердився експериментами Zagotta et al. В цих експериментах рівень відновленої каналами інактивації залежав від концентрації доданого кулькового пептиду. Ефект дії пептиду виявився зворотнім і не спостерігався при попередній обробці пептиду трипсином.
ShakerВ інактиваційна кулька має первинну структуру, що містить 11 гідрофобних або незаряджених залишків та наступну групу з восьми сильно полярних амінокислот. Aldrich та співробітники більш детально дослідили внесок в інактивацію кожної частини кулькового домену. Вони зробили серії селективних мутацій і довели важливість як гідрофобної, так і зарядженої частини домену для процесу інактивації. Модель ShakerВ інактиваційної кульки може бути застосована і до інших типів потенціал-залежнних каналів. Наприклад, було показано, що делеція перших 25 амінокислот NH2 кінцевого домену серцевих К+ каналів щурів призводить до значного зниження швидкої інактивації. Також делеція NH2 кінцевого домену (амінокислот 1-110) RCK4 призводить до експресії повільно інактивованого струму, на відміну від А-типу струму, який виникає внаслідок екпресії RCK4 транскрипту повної довжини. Найбільш визначним є той факт, що ShakerВ кульковий пептид також має здатність блокувати (інактивувати?) еволюційно більш віддалені К+ канали. Було показано, що інші потенціал-залежні Са-активовані калієві канали, і ц-АМФ-залежні канали блокуються ShakerВ кульковим пептидом, якщо його додавати внутрішньоклітинно. Ці спостереження особливо помітні через те, що такі канали за нормальних умов не виявляють інактивації. Дані, отримані з Shaker К+ каналами так само як і з maxi К+ струмами пролили світло на тонкі процеси взаємодії між інактиваційною кулькою та каналом.
Взаємодія кулькового пептиду та рецептора.
Ідея про те, що як гідрофобний так і заряджений кінець кульки роблять свій внесок у взаємодію між кульковим пептидом і рецептором більш інтенсивно досліджувалась шляхом спострежння ефекту мутантних пептидів на інактивацію ShakerВ і maxi К+. В обох випадках було продемонстровано, що зміна загального заряду кульки (внаслідок мутації амінокислот в СООН ділянці) впливає на рівень інактивації як макроструму так і струму через ізольовані окремі канали. Цікаво, що ці зміни заряду пептиду модулюють лише рівень включення інактивації, в той час як рівень виключення інактивації залишається в основному незмінним. Таким чином, збільшення позитивного заряду кульки підвищує рівень включення, і навпаки, зниження позитивного заряду або підвищення негативного зменшує рівень включення інактивації. Ці ефекти електричного заряду максимальні в умовах низької іонної сили і мінімальні при високих сольових концентраціях. Всі разом ці результати було інтерпретовано з точки зору різноманітних елктростатичних взаємодій між позитивно зарядженою кулькою і негативно зарядженою поверхнею в, або поблизу внутрішнього гирла каналу. Електростатичний потенціал, індукований цими негативними зарядами буде спрямовувати кульку до її рецептора. З іншого боку заміна гідрофобних амінокислот полярними і незарядженими залишками з NH2 – кінця кулькового пептиду зменшує рівень макроскопічної інактивації; і цей ефект спостерігається головним чином через збільшення неблокованого рівня. Особливо важливими є мутації, які змінюють гідрофобність в позиції L7 (заміна лейцину). З цих даних видно, що тоді, коли інактиваційний домен взаємодіє зі своїм рецептором, гідрофобні взаємодії зв’язують їх докупи і сила цих взаємодій визначає рівень при якому канали повертаються до неінактивованої конформації, точніше до рівня, коли кулька дисоціює з рецептора.